HDAC6 medeia um mecanismo tipo agressivo para ativação de NLRP3 e inflamassoma de pirina
O MTOC é um “ponto” -tacular
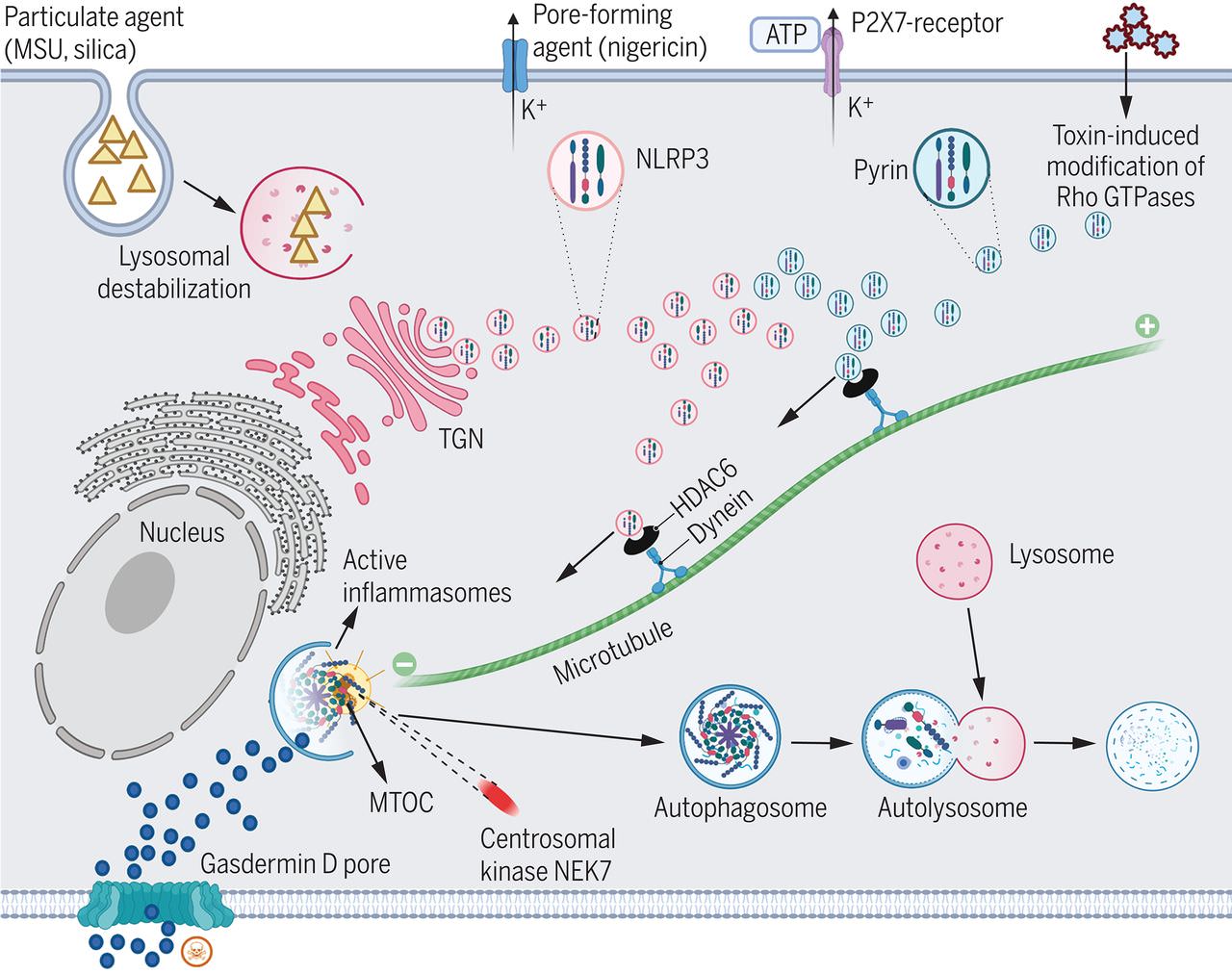
Os complexos inflamassoma são formados em resposta a moléculas associadas a patógenos. Eles iniciam tanto a maturação de citocinas inflamatórias quanto a piroptose, um tipo de morte celular programada. Uma característica notável para a ativação do inflamassoma é a formação de um único ponto supramolecular (ou “ponto”) em cada célula afetada. No entanto, a localização e o mecanismo de formação de manchas são mal compreendidos. Magupalli et al. relatam que para os inflamassomas mediados por NLRP3 e pirina, sua montagem e funções a jusante ocorrem no centro de organização de microtúbulos (MTOC ) Este processo requer o adaptador de dineína HDAC6, que também é um jogador central na formação de agressivos e degradação autofagossômica no MTOC. Este trabalho vincula vários processos celulares importantes e fornece pistas de como os inflamassomas são regulados de forma eficiente.
Ciência , esta edição p. eaas8995
Resumo estruturado
INTRODUÇÃO
RATIONALE
Para decodificar o sítio e a maquinaria molecular na montagem e ativação do inflamassoma em macrófagos, objetivamos visualizar o inflamassoma montagem por imagem celular complementada por inibição farmacológica e deleção direcionada em células e em camundongos.
RESULTADOS
Descobrimos que NLRP3 e inflamassomas de pirina são montados no centrossoma , também conhecido como centro organizador de microtúbulos (MTOC), de cada célula, que serve como o principal local para a ativação da caspase-1 e conversão de IL-1β. Os inibidores farmacológicos da polimerização de microtúbulos, dineína ATPase e o adaptador de dineína histona desacetilase 6 (HDAC6) e deleção direcionada ou knockdown de Hdac6 comprometeram a montagem e ativação desses inflamassomas. Reconstituição de Hdac6
– / –
Para o inflamassoma NLRP3, a localização do MTOC pode facilitar a associação com a quinase localizada no centrossoma NEK7 para aumentar a montagem do inflamassoma. Descobrimos isso em Hdac6 – / –
macrófagos, NLRP3 está preso como pequenas manchas na rede trans-Golgi (TGN ), um local comum previamente reconhecido para associação de NLRP3 após indução por múltiplos estimuladores de NLRP3. Esses dados sugeriram que o transporte de microtúbulos mediado por HDAC6 entrega NLRP3 de TGN para o MTOC. O inflamassoma NLRP3 formado no MTOC colocaliza com LC3b, um marcador de autofagia e um inibidor de autofagia, aumenta a secreção de IL-1β induzida por NLRP3.
CONCLUSÃO
Nosso estudo revela um paralelo inesperado entre a montagem dependente de HDAC6 de NLRP3 e inflamassomas de pirina e a formação de agrossomos no MTOC para degradação autofágica de agregados patológicos ubiquitinados. Os papéis duplos de ativação e inibição desempenhados pela localização do MTOC de NLRP3 e inflamassomas da pirina podem ser críticos para alcançar a regulação equilibrada dos inflamassomas.
Múltiplo estímulos para NLRP3 (MSU, sílica, nigericina e ATP) e pirina (modificação Rho GTPase) são mostrados na parte superior da célula. Os componentes do inflamassoma de NLRP3 e pirina são representados em rosa e azul claro, respectivamente. A montagem do inflamassoma de NLRP3 pode envolver duas etapas: oligomerização inicial no TGN e posterior montagem em um único ponto com a quinase centrossomal NEK7, o adaptador ASC e caspase-1 no MTOC. Este último é obtido após ser transportado retrógrado no microtúbulo pela máquina HDAC6-dineína . A ativação do inflamassoma da pirina também requer HDAC6, dineína e microtúbulos. Os inflamassomas montados no MTOC estão sujeitos à degradação por autofagia de uma maneira s semelhante aos agrossomos que degradam os agregados patológicos. [Figure was created with BioRender (https://BioRender.com).]
“data-hide-link-title=” 0 “data-icon-position=” “href=” https: //science.sciencemag .org / content / sci / 369/6510 / eaas8995 / F1.large.jpg? width=800 & height=600 & carousel=1 “rel=” gallery-fragment-images-1728529576 “title=” HDAC6- e montagem e ativação dependentes de microtúbulos de NLRP3 e inflamassomas de pirina. Estímulos múltiplos para NLRP3 (MSU, sílica, nigericina e ATP) e pirina (modificação Rho GTPase) são mostrados na parte superior da célula. O NLRP3 e os componentes do inflamassoma da pirina são representados em rosa e azul claro, respectivamente. A montagem do inflamassoma NLRP3 pode envolver duas etapas: oligomerização inicial no TGN e posterior montagem em um único ponto com a quinase centrossomal NEK7, o adaptador ASC e caspase-1 no MTOC. Este último é obtido após ser transportado retrógrado no microtúbulo pelo maquinário HDAC6-dineína. A ativação do inflamassoma da pirina também requer HDAC6, dineína e microtúbulos. Os inflamassomas montados no MTOC estão sujeitos à degradação por autofagia de maneira semelhante aos agrossomos que degradam os agregados patológicos. [Figure was created with BioRender (https://BioRender.com).] “> 
HDAC6- e montagem dependente de microtúbulos e ativação de NLRP3 e inflamassomas de pirina.
Vários estímulos para NLRP3 (MSU, sílica, nigericina e ATP) e pirina (modificação Rho GTPase) são mostrado na parte superior da célula. Os componentes do inflamassoma da pirina e do NLRP3 são representados em rosa e azul claro, respectivamente. A montagem do inflamassoma NLRP3 pode envolver duas etapas: oligomerização inicial no TGN e posterior montagem em um único ponto com a quinase centrossomal NEK7, o adaptador ASC e caspase-1 no MTOC. Este último é obtido após ser transportado retrógrado no microtúbulo pela máquina HDAC6-dineína. A ativação do inflamassoma da pirina também requer HDAC6, dineína e microtúbulos. Os inflamassomas montados no MTOC estão sujeitos à degradação por autofagia de maneira semelhante aos agrossomos que degradam os agregados patológicos. [Figure was created with BioRender (https://BioRender.com).]
Resumo
Inflamassomas são complexos supramoleculares que desempenham papéis importantes na vigilância imunológica. Isso é realizado pela ativação de caspases inflamatórias, o que leva à maturação proteolítica da interleucina 1β (IL-1β) e piroptose. Aqui, mostramos que o domínio de ligação de nucleotídeos, repetição rica em leucina e proteína 3 contendo domínio de pirina (NLRP3) – e montagem de inflamassoma mediada por pirina, ativação de caspase e conversão de IL-1β ocorrem no centro de organização de microtúbulos (MTOC ) Além disso, o adaptador de dineína histona desacetilase 6 (HDAC6) é indispensável para o transporte e montagem dos microtúbulos desses inflamassomas tanto in vitro quanto em camundongos. Como o HDAC6 pode transportar agregados patológicos ubiquitinados para o MTOC para formação de agressivos e degradação autofagossômica, seu papel na ativação do NLRP3 e do inflamassoma da pirina também fornece um mecanismo inerente para a regulação negativa desses inflamassomas pela autofagia. Este trabalho sugere um paralelo inesperado entre a formação de agregados fisiológicos e patológicos.

